% P3 t$ l; ~& K7 x# ?& \
2 i3 O) _$ f. J5 ~' X' v" B1 ~% M) S2 C2 y
原创 与癌共舞
* W+ U& G% `; z: L7 e5 J; v) C- ^: U" o) \4 I0 w
3 a: \: }5 a% g) k7 o( `3 O& _
8 ~" n; A, w1 b- [4 c X6 o) T. M/ `. n) U0 I5 k( k
k& U% |# V7 w/ J; X( w8 x' c' B( H% |" ~
( ^' h; s8 h( p) n, F# _0 H
# B' M% a9 p: P4 n2 d2 d) y; g$ w2 L2 D p1 a! H
# @0 N% ^; q; v! J3 R+ x$ w$ [( n
7 ~4 }9 ^7 A) @) c2 S+ ?! B; |
7 `" M! Y8 q, r8 Q( k近日,瑞士洛桑大学的研究小组取得一项新成果。他们发现阻断CD36后,肿瘤内的浸润的调节性T细胞减少,杀伤性T细胞则明显增加,从而抑制肿瘤的生长,同时免疫稳态并没有被破坏。不但如此,阻断CD36还可以和免疫治疗联合使用,进一步增强对肿瘤的杀伤。相关研究发表在了《自然·免疫学》杂志上。
; b% X2 M9 R0 a/ y8 m( g* O4 `* k, n$ R
9 M" d* c$ ~1 \- R
6 C+ J/ a# u' S/ ]3 G0 r
5 B* u6 Z% |/ k1 V% R% d" }2 {' W1 ^; Y; o0 O3 I
" M R: }7 b5 f
在免疫系统中,杀伤性T细胞和调节性T细胞是一对背靠背的好搭档,在杀伤性T细胞对病原体和异常细胞大开杀戒的同时,是调节性T细胞保障它不会伤害健康组织。
* x4 t9 Q1 N a7 c8 Z+ @
$ y" G* x. [1 N我们知道,在恶性肿瘤中,肿瘤免疫抑制是肿瘤微环境的最重要特征之一,通过免疫抑制微环境,肿瘤细胞可以逃避机体免疫系统的识别和攻击,产生免疫逃逸。既往研究发现,杀伤性T细胞在肿瘤中的浸润会激活一些免疫调节机制,把调节性T细胞也招募到肿瘤中。这些调节性T细胞就会产生“叛变”,抑制了杀伤性T细胞杀灭肿瘤细胞。
3 _9 g, \, V; G# `如果能靶向性地抑制肿瘤中的调节性T细胞,是不是能让杀伤性T细胞提高抗肿瘤免疫能力,同时又不影响正常的免疫调节功能?
, K+ z( _" `+ o" ~3 }- E$ T$ T2 h
3 p7 I, j( G; @: v. [" H5 w3 {+ v
为了探索这个问题,首先就要知道调节性T细胞是如何在肿瘤微环境中安定地生存的。瑞士洛桑大学研究人员首先分析了一些乳腺癌患者的调节性T细胞的基因表达特征,并且比较肿瘤中浸润的和外周循环中的调节性T细胞的差异。
( c9 ?7 e* M! }9 w
: j/ Y* v) n0 Q t9 G
他们发现,这两类调节性T细胞的参与脂质代谢的基因表达差异很明显,其中就包括CD36。CD36是一种脂肪酸受体,负责脂肪酸的识别和细胞跨膜转运,CD36能帮助肿瘤细胞从周围环境中摄取脂肪分子,这些脂肪分子成为这些细胞转移的能量来源。在肿瘤浸润的调节性T细胞中,CD36的表达水平更高。
& s/ n6 K' j- h3 n" y. O
% H+ {+ w2 H: [' x; m
) Y/ J0 _$ M9 f& G
5 z- n5 P# Y) j) j. H
" r, e( i; h( H; _3 C: B: H5 ]肿瘤内调节性T细胞脂质积聚及CD36表达增加
; R' ~8 C6 Y% I* s# y不仅仅是乳腺癌患者,研究人员在人肺癌细胞和黑色素瘤细胞体外模型,以及黑色素瘤和肠癌小鼠模型中都发现了这一现象。
# w6 l2 }6 r2 Z4 ]* R" \
在多种肿瘤模型中都发现了这一现象,说明这并不是一个偶然的事件,而是普遍存在的。接下来,研究人员对CD36表达水平的影响进行了研究。
: g$ ?; o- @' C) }1 P2 c% ?他们构建了调节性T细胞特异性CD36缺陷型小鼠,来观察缺乏CD36是否会对免疫系统产生破坏。结果发现,缺少CD36也不会影响T细胞激活标志物,以及调节性T细胞的标志性分子Foxp3的表达。CD36对免疫系统稳态的维持不是必需的。
: T7 l. S2 J0 b, a/ U
而肿瘤内促炎细胞因子干扰素-γ(INF-γ)和肿瘤坏死因子(TNF)的水平明显增加,这意味着,CD36的存在阻碍了CD36抑制肿瘤内调节性T细胞产生促炎细胞因子的能力。
( B' P! g' o9 {' X除此之外,CD36的缺失不但影响了肿瘤内调节性T细胞的功能,还减少了它们在肿瘤中的聚集。荧光染色实验结果显示,CD36缺失并没有影响肿瘤内调节性T细胞的增殖,但是促进了细胞的凋亡。既往研究发现,线粒体与调节性T细胞的抑制功能和生存密切相关。
( Q; X( Q. ?6 s" \( r
研究人员推测,肿瘤内调节性T细胞的CD36表达增加可能是通过调节线粒体适应性来响应肿瘤微环境,改变了代谢途径,帮助调节性T细胞更好地生存。随后的实验证明了他们的推测。
" @7 |/ G2 ?5 q6 P5 Z
那么CD36是通过谁来调节线粒体代谢适应性的改变的呢?这究竟是哪个基因进行调控的呢?通过对比发现,肿瘤内调节性T细胞的参与PARP通路的基因表达增加,而CD36可以通过线粒体激活因子PARP-β和PARP-γ依赖性的方式增强对线粒体活性的调节,来改变代谢。
1 H9 i- D/ h( P' W2 ?9 F经过逐一实验,研究人员将目标锁定在了PARP-β上。PARP-β途径的激活使得肿瘤内调节性T细胞线粒体膜电位增加,凋亡减少,在不缺失CD36的情况下,PARP-β的激活还增强了CD36的表达,进一步提高了肿瘤内调节性T细胞的代谢适应。
/ P5 [: G2 @: ]( r, C, l: I) ~# N U- U
& V) h6 W! _0 `- y# d
9 m) L. k5 O" P% G0 @+ n: p, z: @1 y9 M/ C; d
/ ]& o# T: \# p) p' v
最后,为了验证抗癌效果,研究人员制作了靶向CD36的单克隆抗体,并对黑色素瘤小鼠进行了治疗。结果发现,肿瘤中调节性T细胞的水平降低,杀伤性T细胞的浸润增加,显著抑制了肿瘤的生长。
4 n, S K* J! d4 \. L5 g; k3 }
然而,肿瘤中杀伤性T细胞的耗竭也是常常出现的问题。单独抑制调节性T细胞可能是不够的,还需要一些额外的“助力”。研究人员尝试将CD36单克隆抗体和免疫治疗结合起来,同时使用阻断CD36和PD-1。
/ g1 L) v) T E4 M0 V' D
他们将小鼠分为四个小组:第一组是普通肿瘤小鼠(蓝),第二组是调节性T细胞CD36特异性缺失小鼠(红),第三组接受PD-1治疗的小鼠(黑),第四组为同时阻断CD36和PD-1治疗的小鼠(绿)。
8 V& K/ q, X; X结果发现,与其他三组相比,联合治疗对肿瘤细胞的杀伤力更为强劲,也延长了小鼠的生存期,超过50%的小鼠活过了28天。
& S- P$ [4 }2 U) E+ W
& X5 I) l T: L" c8 t* W
+ J0 G7 \. D2 y8 E( S4 \
* f$ z; D1 |; V" Q! V* F
, f* R" s( e: ]6 e$ x$ @
+ |( j$ @1 U9 T! t E: p) k& p/ f
这项研究表明,CD36让肿瘤中的调节性T细胞在代谢方面异于其他调节性T细胞,使它们能更好地适应肿瘤微环境,抑制了杀伤性T细胞杀灭肿瘤细胞。靶向抑制CD36,则会令肿瘤中的调节性T细胞“弃暗投明”,这或许可以成为新的抗癌疗法或是辅助抗癌疗法。
% Q5 O, E% {& N# B
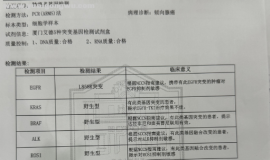
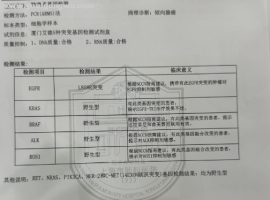
 奥西替尼耐药后续治疗,出现腹水
治疗经过:
2010确诊腺癌
治疗方案:右肺上叶切除,10年民诺宾2次,顺铂+盖诺2次。
奥西替尼耐药后续治疗,出现腹水
治疗经过:
2010确诊腺癌
治疗方案:右肺上叶切除,10年民诺宾2次,顺铂+盖诺2次。
 三代伏美换一代特罗凯获益率问题
家父25年2月因肩膀疼痛确诊肺腺癌晚期4B,双肺转,多发骨转,脑转(非典型),
三代伏美换一代特罗凯获益率问题
家父25年2月因肩膀疼痛确诊肺腺癌晚期4B,双肺转,多发骨转,脑转(非典型),
 盲试靶向药 29个月,治疗分享
时间:2025/1/27 盲试靶向药第29个月。
本月治疗方案:7080(14mg)。2024年11月底
盲试靶向药 29个月,治疗分享
时间:2025/1/27 盲试靶向药第29个月。
本月治疗方案:7080(14mg)。2024年11月底
 完成手术,病理也出来了,烦请各位老
之前因为心脏问题,去心内造影,前降支75%狭窄,回到心胸外科手术,目前主治医生说是
完成手术,病理也出来了,烦请各位老
之前因为心脏问题,去心内造影,前降支75%狭窄,回到心胸外科手术,目前主治医生说是
 生存期超4年!首个击败奥希替尼的药
作者:seacat
2025年3月26日,巴黎举行的2025欧洲肺癌大会(ELCC 2025)报道了III期研
生存期超4年!首个击败奥希替尼的药
作者:seacat
2025年3月26日,巴黎举行的2025欧洲肺癌大会(ELCC 2025)报道了III期研


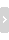


 3 a: |0 j: |9 d8 {' N
3 a: |0 j: |9 d8 {' N ! y: Z E% p- x1 ^- [
! y: Z E% p- x1 ^- [ ) J! g9 |9 @. H0 A m1 u
) J! g9 |9 @. H0 A m1 u


 9 n# W3 H6 j- E* Z4 M, U+ I
9 n# W3 H6 j- E* Z4 M, U+ I
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡