| |
| |
| |
| |
 23年脑膜转,目前脑部稳定,肺原发灶
2025.3至2025.5.22脑实质和脑膜稳定,肺部原发灶及胸腔积液控制不住。CEA持续升高。
23年脑膜转,目前脑部稳定,肺原发灶
2025.3至2025.5.22脑实质和脑膜稳定,肺部原发灶及胸腔积液控制不住。CEA持续升高。
 林根:小细胞肺癌放疗及免疫治疗常见
整理者:雨过天晴审核人:鹰版小细胞肺癌(SCLC)是一种恶性程度高、侵袭性强的肺癌亚
林根:小细胞肺癌放疗及免疫治疗常见
整理者:雨过天晴审核人:鹰版小细胞肺癌(SCLC)是一种恶性程度高、侵袭性强的肺癌亚
 求助:肺腺癌3年,脑转脑膜转骨转,E
肺腺癌3年,脑转脑膜转、骨转,E19+797S突变,免疫阴性,阿美29个月,期间放疗2次,后
求助:肺腺癌3年,脑转脑膜转骨转,E
肺腺癌3年,脑转脑膜转、骨转,E19+797S突变,免疫阴性,阿美29个月,期间放疗2次,后
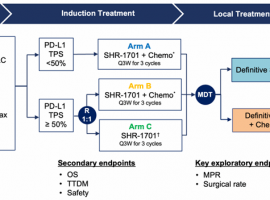 25%肺癌患者变不可手术为可手术,新
作者:seacat
驱动基因阴性的III期非小细胞肺癌的治疗颇具挑战,能否手术切除没有一致
25%肺癌患者变不可手术为可手术,新
作者:seacat
驱动基因阴性的III期非小细胞肺癌的治疗颇具挑战,能否手术切除没有一致
 聚焦2025 ASCO 前列腺癌新进展,从循
作者:Tony男人一旦上了年纪,总有些难以启齿的尴尬,如尿频、尿急、尿不尽……很多人
聚焦2025 ASCO 前列腺癌新进展,从循
作者:Tony男人一旦上了年纪,总有些难以启齿的尴尬,如尿频、尿急、尿不尽……很多人




 显身卡
显身卡


